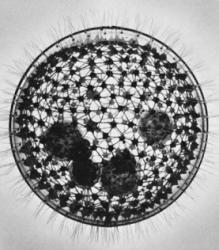
Zelené riasy rodu Volvox (slovensky váľače, česky váleče) vytvárajú guľovité kolónie v ktorých sú tisíce buniek uzavreté do priehľadnej gule tvorenej želatíne podobným glykoproteínom. Na základe predchádzajúcich výskumov zo 70tych rokov minulého storočia sa zrodilo presvedčenie, že prvé takéto organizované spoločenstvá – predchodcovia váľačov – sa na Zemi objavili niekedy pred 50 až 75 miliónmi rokov. Najnovšie výskumy, využívajúce moderné technické možnosti rýchlej sekvenácie DNA, porovnanie genetických odlišností medzi jednotlivými druhmi a porovnanie s dobre datovanými fosíliami však tento časový úsek predlžujú na troj- až štvornásobok. Podľa článku, ktorý zverejnilo najnovšie číslo amerického časopisu PNAS (Proceedings of the National Academy of Sciences) sa jednotlivé jednobunkové riasy „odhodlali“ na spoluprácu v rámci mnohobunkovej kolónie a vytvorili tak prvého evolučného predka dnešných vaľačov niekedy pred minimálne 200 miliónmi rokov, počas triasu. Bolo to obdobie teplej vlhkej klímy, doba stromových papradí a dinosaurov.
Vedci pod vedením Matthewa D. Herrona z Arizonskej univerzity skúmali sekvencie DNA 51 rôznych druhov mnohobunkových organizmov, prevažne rias z rodu váľačov (Volvox) a im príbuzných druhov a snažili sa zrekonštruovať vývojový rodostrom čeľade Volvocaceae. V tejto zaujímavej skupine rias sú druhy, ktoré vytvárajú jednoduché zoskupenia štyroch identických buniek, iné sa podobajú guľatému zhluku 32 až 64 nie úplne rovnakých buniek, kde už niektoré majú špecializované funkcie, ale na druhej strane sem patria druhy s 50timi tisícami špecializovaných buniek. Aká je teda odpoveď na otázku, kedy sa na Zemi objavil ich prvý spoločný predok, prvá forma takejto kolónie kooperujúcich buniek? Pred asi 223 miliónmi rokov sa začali jednotlivé bunky zhlukovať a obaľovať sa do výlučkov z cukrov a bielkovín. O asi 20 miliónov rokov neskôr sa v takomto spoločenstve vyvinula geneticky riadená kontrola delenia podľa toho, ako vhodné sú podmienky prostredia. To bol prvý krok na ceste k plánovanej stavbe tela organizmu s geneticky kontrolovaným limitom pre veľkosť. Až asi o desať miliónov neskôr si spoločenstvo začalo koordinovať smerovanie bunkových bičíkov smerom von. Vznik pohlavných zárodočných buniek sa podľa výskumov datuje do obdobia pred asi 100 miliónmi rokov.
Odhaliť podrobnosti vývojového kroku, ktorý viedol k jednoduchým formám spolupráce a následnej prvotnej špecializácie medzi jednotlivými bunkami v rámci uzavretej biologickej štruktúry pomáha pochopiť aj vývojovú cestu vzniku organizovaných mnohobunkových organizmov ako sú rastliny a živočíchy. Rozmanité, evolučne úspešné symbiotické formy spolužitia pôvodne samostatných buniek sa na Zemi počas biologickej evolúcie vyvinuli mnohokrát a podnecovali rozvoj rôznorodosti zložitejších mnohobunkových organizmov. Spätný pohľad na vývoj každého, aj toho najkomplikovanejšieho organizmu vedie k jednobunkovým formám života, ktoré sa na Zemi vyvíjali po viac ako 2 miliardy rokov. Až na určitom stupni vývoja vzniká tlak vzájomnej konkurencie, potreba ochrany a obrany, čo zvýhodňuje vytváranie zoskupení s určitým prerozdelením životných funkcií. Jednobunkový organizmus tým získava a stráca zároveň.
Dve kľúčové inovácie, vyplývajúce z ranného štádia spolupráce, konkurenčného súboja a následného zmieru medzi jednotlivými bunkami viedli k rýchlej integrácii a k rozšíreniu mnohobunkových foriem v skupine váľačov. Keď bunky vytvoria spoločnú štruktúru, môže ich to ochrániť pred predátormi, pre ktorých je takéto sústo už priveľké. Každé spolužitie však niečo stojí a budovanie vonkajšej medzibunkovej konštrukcie a produkcia glykoproteínov na tvorbu spoločného domova, ktorý chráni kolóniu je daňou za jednotnú, v prípade váľačov nie kruhovú, ale doslova guľovú obranu. Ďalším prispôsobením je potreba podriadiť sa nejakej deľbe práce a kompetencií, čo znamená, že nie každá bunka bude mať právo na reprodukciu. A to predstavuje zdroj zásadného konfliktu záujmov jednotlivých buniek. Jeho prekonanie je asi tým najvýznamnejším krokom k vývoju mnohobunkových organizmov. Ako rozmnožovanie vyriešili váľače, to vytrvalému čitateľovi ponúka záver tohto článku. Pri nepohlavnom spôsobe rozmnožovania kolónie, v ktorých len niektoré bunky majú právo sa deliť, je nevyhnutné sa vyhnúť akémusi „podvádzaniu“ a zaviesť mechanizmy „kontroly pôrodnosti“, podobne ako v Číne. To všetko sa musí vyriešiť už na tej najjednoduchšej úrovni spolupráce. Ako sa zmier v týchto konfliktov prejavil v genetickom zázname a či veľkosť kolónie ovplyvňuje mieru diferenciácie (stupeň „deľby práce“) buniek – to sú ďalšie ciele pokračujúceho výskumu Matthewa Herrona a jeho kolegov.
Kredit: American Museum of Natural History, New York City
Dnešné váľače (Volvox) sú zelené fotosyntetizujúce riasy, ktoré vytvárajú takzvané cenóbie. Sú to zvláštne duté gule zo želatínovitého glykoproteínu, v ktorom sa nachádzajú tisíce bičíkatých buniek, poprepájaných jemnými vláknami z cytoplazmy. To udržuje stabilnú štruktúru a formu cenóbie a umožňuje celej kolónii sa koordinovane pohybovať vďaka bunkovým bičíkom, ktoré smerujú von z povrchu. Bunky sa však museli evolučne podriadiť aj ďalším formám spolupráce a špecializácie. Pre zjednodušenie vysvetlenia prirovnajme cenóbiu k zemeguli. Tiež má akési dva póly a rovinu rovníka, kde bunky majú v rámci deľby práce iné poslanie. Na jednom z pólov, teda na jednej strane guľovej cenóbie majú bunky vyvinutejšie svetlocitlivé škvrny, čo pomáha celej kolónii určiť odkiaľ prichádza svetlo a pomaly sa tým smerom pohybovať. Táto fototaxia sa dá pozorovať aj pod mikroskopom. Ak kolónie rias budú mať dosť priestoru pod krycím sklíčkom, budú sa snažiť sa presúvať smerom k osvetleniu.
Váľače totiž potrebujú pre fotosyntézu dostatok svetla. Preto ich domovom sú presvetlené, dostatočne čisté a teplé stojaté prírodné vody. Najčastejšie sa v nich nachádzajú dva druhy – menší váľač zlatý Volvox aureus, ktorý sa pri veľkosti pol milimetra javí aj pre pozorné skúmajúce oko len ako zelená bodka. Jednotlivci – jednobunkové riasy sú však väčšie (5 až 8 mikromerov), no je ich menej, než pri druhu váľač gúľavý (váleč koulivý Volvox globator). Jeho guľaté kolónie s väčším počtom menších, 4 mikrometre veľkých jedincov dosahujú až 2 milimetre.
To čo je však na váľačoch zaujímavé viac, ako tvar kolónií a ich pohyb, je ich rozmnožovanie. Svojich potomkov cenóbie produkujú nepohlavným aj pohlavným spôsobom. Mikroskop objaví vo vnútri väčšiny guľových kolónii ich akoby miniatúrnejšie kópie – dcérske kolónie. Sú výsledkom nepohlavnej reprodukcie buniek, ktoré sa nachádzajú v rovine uprostred medzi oboma pólmi cenóbie – akoby v rovine „rovníka“ celej gule. Ich dôležitou úlohou je deliť sa a deliť sa, až sami vytvoria miniatúrne nové kolónie. Keď „dospievajú“, musia sa prevrátiť naruby, aby bičíky, ktoré smerujú spočiatku dovnútra sa obrátili smerom von. Na slobodu sa prebojujú až pri rozpade materskej kolónie.
V oblasti „rovníka“ sú však ďalšie špecializované, takzvané zárodočné bunky. Sú dvoch druhov – samčie, ktoré delením tvoria spermie a samičie, ktoré sa nedelia, len sa pretransformujú na vajíčko. Väčšina kolónií je obdarená samičími aj samčími zárodočnými bunkami. Spermie a vajíčka sa však vyvíjajú a do vonkajšieho prostredia uvoľňujú v rôznom čase a tak váľač zabraňuje samooplodneniu. Oplodnené vajíčko – zygota – podľa ktorej sa dá pomerne ľahko určiť druh válača, si vytvára pevný ochranný obal. Vďaka nemu dokáže prežiť neľútostné podmienky zimy a v nasledujúcom lete zabezpečiť znovuzrodenie týchto zaujímavých rias predstavujúcich ranné štádium prechodu od jednobunkovej k mnohobunkovej forme života.

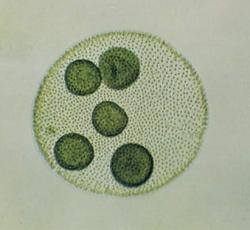
Guľovité cenóbie váľača gúľavého (Volvox globator). Zelené gulôčky sú dcérske kolónie, ktoré sa vznikajú vo vnútri dutých kolónii nepohlavným delením určitých buniek.
Nasledujúce snímky sú len odkazmi na stránku Wima van Egmonda, kde nájdete obrázky v plnom rozlíšení. Vysvetľujúce texty však ponúkame v preklade.
 |
Detail jednotlivých buniek na povrchu dutej guľovitej cenóbie. Dva bičíky každej bunky smerujú von a zabezpečujú pohyb celej kolónie. Červené bodky na každej bunke sú svetlocitlivé škvrny, ktoré umožňujú vnímať intenzitu svetla. Na jednej strane má kolónia bunky, ktoré tieto bunkvé organely má vyvinuté viac. Obrázky v plnom rozlíšení – stránka Wima van Egmonda |
 |
Cytoplazmatické vlákna spájajú jednotlivé bunky a pomáhajú udržiavať stabilnú priestorovú štruktúru cenóbie. Obrázky v plnom rozlíšení – stránka Wima van Egmonda |
 |
Dcérska kolónia, ktorá vznikne nepohlavným delením špecializovanej bunky vo vnútri kolónie sa musí pred dokončením vývoja pretočiť naruby, aby bičíky, ktoré smerovali dnu sa otočili smerom von. Obrázky v plnom rozlíšení – stránka Wima van Egmonda |
 |
„Dospelé“ dcérske kolónie sa zo zajatia materskej cenóbie uvoľnia až keď sa rozpadne. Obrázky v plnom rozlíšení – stránka Wima van Egmonda |
 |
Kolónia uvoľňuje do prostredia balíčky spermií. Obrázky v plnom rozlíšení – stránka Wima van Egmonda |
 |
Oplodnené vajíčka – zygóty – zapuzdrené v ochrannom obale, ktorý umožňuje prežiť nepriaznivé obdobia. Napríklad zimu, či sucho. Obrázky v plnom rozlíšení – stránka Wima van Egmonda |
 |
Kolónie váľačov majú aj doslova vnútorných predátorov – napríklad vírniky, ktoré sa prevŕtajú cez stenu dnu, kde sa pasú na jednotlivých bunkách rias. Pri nebezpečenstve, keď ustane pohyb kolónie, vírnik rýchlo kolóniu opustí. Obrázky v plnom rozlíšení – stránka Wima van Egmonda |
Prameny:
Science Daily
PNAS
Stránka Wima van Egmonda
http://www.microscopy-uk.org.uk/mag/indexmag.html?http://www.microscopy-uk.org.uk/mag/artdec03/volvox.html
Diskuze:


